Intellau_Celistic
5'3 KHHV Mentalcel
★★★★★
- Joined
- Aug 26, 2021
- Posts
- 165,059
Empathizing-Systemizing Theory suggests that low empathizing and high systemizing are linked to autistic traits in the general population. Evidence from autistic individuals is convincing, but more research in the normal population is needed. Method: We conducted two surveys (N=3,345) investigating the relationships between empathizing, systemizing and autistic traits in the general population, using a large variety of self-report instruments and direct performance tests. Results: Strong connections between autistic symptoms, empathizing, and systemizing were found using commonly used measures (Autism Quotient, Systemizing Quotient and Empathizing Quotient). Other measures on empathizing and systemizing found the connections that E-S-theory predicts, but the correlations were a lot more modest. Weak empathizing was related to autism's social difficulties, while systemizing was linked to non-social aspects of autism. Conclusions: The present results support the main tenets of empathizing-systemizing theory, but suggest that earlier findings might be inflated due to overlapping items in the most common assessment instruments.
Introduction
In this paper three main themes are considered. First, I analyse the importance of the present moment in family therapy. Second, I explore the polyphony of voices as the main aspect of human psychology and its meaning for family therapy dialogue. Third, the effectiveness of dialogism in the treatment of psychotic problems will be illustrated in the province of Western Lapland in Finland.
The present moment
Open dialogue describes both organizing the psychiatric practice for severe crises and dialogues in meetings with the family and the rest of the client's social network. For therapists the main challenge becomes being present and responding to every utterance. We are living in the ‘once occurring participation in being’ (Bakhtin, 1993).
Tom Andersen (2007) was preoccupied by three different realities of our practices as clinicians. In the ‘either–or’ reality we handle issues that are visible but dead in the sense that they are exactly defined and the definitions remain the same in spite of the context. In the ‘both–and’ reality we deal with issues for which many simultaneous descriptions are possible. These issues are living and visible. This is the case, for instance, in the family therapy discussion when we make space for different voices to become heard without considering one point of view to be right and another wrong. The most interesting may be the ‘neither–nor’ reality, in which things are invisible but living. We experience something as taking place, but we do not have an exact linguistic description for it. We may say that it is neither this nor that, but I know that something is taking place. As an example, Andersen gives handshaking. It is something that happens in our embodied participation in the session, yet it is not commented on by words but remains as our embodied experience of the present moment.
Daniel Stern (2004) in emphasizing the importance of the present moment is critical of descriptions of psychotherapy and psychoanalysis that focus on clients' narratives. The therapist is seen as the one giving meanings to patients' stories, in different schools in different ways. Therapy deals with explicit knowledge in linguistic descriptions. Stern proposes moving from explicit knowledge to the implicit knowing that happens in the present moment as embodied experience, and mainly without words. We live in it. The present moment is a short one, varying between one and ten seconds, lasting on average three seconds.
Stern is describing individual psychotherapy. In the type of family therapy that focuses on generating dialogues this means shifting the focus from the content of narratives to the present moment when narratives are told. Therapists and clients live in a joint embodied experience that happens before the client's experiences are formulated in words. As in dialogue on the whole an intersubjective consciousness emerges. Our social identity is constructed by adapting our actions to those of others. For Bakhtin, knowing myself is only possible by seeing ourselves through the eyes of the other. I see myself through others' eyes (Bakhtin, 1990). In Bakhtin's view if we want to see ourselves as living persons while looking at our reflection in the mirror, we adapt others' eyes to do that. Living persons emerge in real contact with each other and adapt to each other as in a continuous dance in automatic movements without controlling their behaviour in words.
Intersubjectivity
The intersubjective quality of our consciousness is shown in the mother–baby communication studies conducted by Colwyn Trevarthen (1990). Trevarthen's careful observations of parents and infants demonstrate that the original human experience of dialogue emerges in the first few weeks of life, as parent and child engage in an exquisite dance of mutual emotional attunement by means of facial expressions, hand gestures and tones of vocalization. This is truly a dialogue: the child's actions influence the emotional states of the adult, and the adult, by engaging, stimulating and soothing, influences the emotional states of the child. Bråten (1992, 2007) describes the Virtual Other as an innate part of the baby's mind that in a way waits for a dialogue with the Actual Other. If the Actual Other is not present, the dialogue emerges with the Virtual Other. Near relations take place in the mode of felt immediacy, in feelings that are felt in a pre-linguistic form. David Trimble has been analysing the importance of Trevarthen's and Bråten's studies for family and network interventions (Seikkula and Trimble, 2005).
In every meeting two histories happen. The first is the history generated by our presence as embodied living persons. We adapt ourselves to each other and create a multi-voiced polyphonic experience of the shared incident. Salgado and Hermans (2005) point out that we cannot call this ‘experience’, because experience already presumes psychological meaning that is included by the Other or Otherness in the situation. It is our embodied experience for which manifold meanings emerge based on the number of participants in the situation. Family sessions as such already include several family members and often two or three therapists. Most of this history takes place without words, but not all. The words that refer to our presence in this conversation often include the most important emotions connected to those voices of our lives that deal with difficult experiences. We may, for example, describe and reflect on our feelings about the specific situation we are talking about.
The second history in the same situation occurs in the stories that living persons tell of their life. Stories always refer to the past, they never can reach the very present moment, since when the word is formulated, and when it becomes heard, the situation in which it was formulated has already passed. Integrating the two aspects of the same moment it becomes evident what focus on dialogue can add into narrative orientation. While comparing narrative and dialogical approaches in family therapy, Roger Lowe (2005, p.70) stated that:
Malignant mesothelioma is a relatively rare cancer arising from the mesothelial lining of pleura as well as the peritoneal cavities, tunica vaginalis, and pericardium. Most (∼80%) cases arise from the pleural mesothelium, and of these, most (∼60–70%) are associated with asbestos exposure.1 Several studies have reported on the epidemiology of mesothelioma from patients registered in the United States National Cancer Institute's Surveillance, Epidemiology, and End Results (SEER) Program database.1, 2, 3, 4, 5 There are presently ∼2500 to 3000 malignant mesothelioma cases per year, mostly among older, white males; the United States incidence seems to have peaked in the years 2000–2005. The recent decline in incidence is attributable to declining asbestos exposure, and this trend is expected to continue.1
Pleural mesothelioma is an often debilitating malignancy with a very poor prognosis, in part because malignant mesothelioma is often diagnosed when patients become symptomatic with advanced-stage disease. For untreated disease, the median survival (MS) is generally less than a year. Treatment options generally include palliative surgical resection, radiation therapy, chemotherapy, and/or pleurodesis, which can palliate symptoms of pain and respiratory decline, delay symptomatic progression, and/or prolong survival. Radical, curative-intent multimodality therapy, with extrapleural pneumonectomy, adjuvant hemithoracic radiation therapy, and adjuvant chemotherapy (referred to as trimodality therapy), can be offered to select few patients with localized disease and adequate pulmonary and cardiac function.6, 7 In these patients, epithelioid histology (versus sarcomatoid or mixed histologies), no regional lymph node involvement, and negative surgical margins are favorable predictors of survival outcome.8, 9 Among patients with favorable prognostic factors, the MS can be on the order of 4 to 5 years.8, 10
This study offers a descriptive, retrospective analysis of patients with malignant pleural mesothelioma registered in the population-based SEER database. This study was undertaken with the goal of better characterizing the overall survival (OS) of malignant pleural mesothelioma and the risk factors affecting OS. Recently, there have been several Italian population-based studies of survival among patients with malignant pleural mesothelioma.11, 12, 13 To our knowledge, this is the first article in over 20 years14 to analyze the OS of patients registered in the SEER database with a diagnosis of malignant pleural mesothelioma, and it currently represents the largest survival analysis of patients with malignant pleural mesothelioma.
The incidence of malignant pleural mesothelioma (MPM) is higher in men than in women, likely due to increased occupational asbestos exposure among men. Women also appear to experience better long-term survival. This study evaluates the role of gender in relation to established prognostic factors in MPM.
Methods
We reviewed 715 cases of MPM treated with extrapleural pneumonectomy at our institution between July 1987 and December 2008. Data for patients with epithelial and nonepithelial tumors were analyzed separately. Kaplan-Meier and Cox regression analyses were used to estimate survival for various cohorts to assess the relationship between gender and survival independent of age at surgery, stage, side, and preoperative laboratory studies.
Results
Of the 702 patients with complete data available, 114 out of 450 patients with epithelial tumors and 31 out of 252 patients with nonepithelial histology were women. Women with epithelial (and not nonepithelial) disease were found to differ significantly from men with respect to younger age, higher rate of thrombocytosis, and longer survival after surgery. The effect of gender on survival of patients with epithelial disease persisted when controlling for age, stage, thrombocytosis, leukocytosis, and anemia with a multivariable analysis. No significant differences in survival were seen among patients with nonepithelial disease with regard to gender, age, or anemia.
Conclusions
In the absence of other negative prognostic factors, women with epithelial MPM demonstrated a survival advantage. These findings support an aggressive approach to treating MPM including extrapleural pneumonectomy in individuals with favorable prognostic predictors, particularly women with epithelial histology and no other risk factors.
Malignant pleural mesothelioma (MPM) is a rare and aggressive disease arising from the pleural mesothelium, with a reported survival of less than 12 months.1 Despite advances in modern systemic chemotherapy, long-term survival in patients with MPM remains limited.2 Because of the lag time between asbestos exposure and disease manifestation, the peak incidence of MPM in the United States is projected to be between 2010 and 2020. Other developed countries show similar epidemiological trends, and the health, medicolegal, and industrial implications of this disease will continue to intensify in the years to come.3, 4, 5
The standard of care for patients with MPM has not been established. Extrapleural pneumonectomy (EPP) has been performed as a treatment option.6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63 This procedure involves en bloc resection of the parietal pleurae, lung, ipsilateral hemidiaphragm, and ipsilateral pericardium. A number of studies have demonstrated encouraging results for selected patients with MPM who underwent EPP. However, because of the relentless nature of the disease, treatment failure after EPP alone remains high.48 Since then, a number of institutions have combined EPP with adjuvant chemotherapy and/or radiotherapy. In comparison with procedures such as pleurectomy and decortication, EPP aims to achieve radical cytoreduction and facilitate maximal delivery of postoperative radiotherapy.
Despite a heightened interest in EPP over the past decade, concerns about the morbidity and mortality of this surgical procedure, and its efficacy, have delayed a consensus in its practice. In addition, there is a lack of robust clinical data on prognostic factors for overall survival and quality of life evaluation. To date, no randomized controlled trials (RCTs) have been conducted to examine the potential benefits of EPP. The most recent systematic review conducted by Maziak et al.64 provided an informative summary of existing literature up to February 2004. However, significant changes have taken place since that time. More recently, innovative multimodalities such as neoadjuvant chemotherapy and hyperthermic intraoperative chemotherapy (HIOC) have been evaluated.6, 7, 8 The current systematic review aims to determine the efficacy of EPP either alone or as part of a multimodality therapy in the current medical setting, with a primary focus on survival and perioperative outcomes. In addition, quality of life assessments were also systematically examined.
METHODS
Literature Search Strategy
Electronic searches were performed using Ovid Medline, EMBASE, Cochrane Central Register of Controlled Trials, Cochrane Database of Systematic Reviews, and Database of Abstracts of Review of Effectiveness from January 1985 to January 2010. To achieve the maximum sensitivity of the search strategy and identify all studies, we combined “mesothelioma” as a Medical Subject Headings (MeSH) term or a keyword and “pneumonectomy” as a MeSH term or keyword. The reference lists of all retrieved articles were reviewed for further identification of potentially relevant studies. All relevant articles identified were assessed with application of inclusion and exclusion criteria.
Selection Criteria
Eligible studies for the present systematic review included those in which patients with histologically proven MPM were treated by EPP. Adjuvant therapy included chemotherapy, radiotherapy, photodynamic therapy, and hyperthermic or normothermic intrapleural chemotherapy. Neoadjuvant therapy included systemic chemotherapy. For studies that included patients who underwent EPP as a subset of patients who had other treatments, results for patients who underwent EPP were extracted when possible. When centers have published duplicate trials with accumulating numbers of patients or increased lengths of follow-up, only the most complete reports were included for qualitative appraisal. It is acknowledged that criteria for patient selection for EPP varied among institutions and sometimes within an institution in different time periods. All publications were limited to human subjects and in English language.
Abstracts, case reports, conference presentations, editorials, and expert opinions were excluded. Review articles are omitted due to potential publication bias and possible duplication of results. Studies published before 1990 and those that included 10 or fewer patients who underwent EPP were also excluded.
Data Extraction and Critical Appraisal
Findings from initial scoping searches were used to decide outcomes for the present review. The primary outcomes included overall survival, 30-day mortality and morbidity, and quality of life assessments. The secondary outcomes included disease-free survival, recurrence rates, prognostic factors on overall survival, intraoperative blood loss, length of hospitalization, and operation time. All data were extracted from article texts, tables, and figures. Two investigators (C.Q.C. and T.D.Y.) independently reviewed each retrieved article. Discrepancies between the two reviewers were resolved by discussion and consensus. The final results were reviewed by the senior investigators (P.G.B. and B.C.M.).
RESULTS
Quantity of Trials
A total of 428 references were identified through the five electronic database searches. After exclusion of duplicate or irrelevant references, 121 potentially relevant articles were retrieved for more detailed evaluation. After applying the selection criteria, 58 remained for assessment (Table 1). A number of centers published studies with duplicating patients at different follow-up periods or different primary objectives. Thirty-four of 58 studies from 26 institutions containing the most complete or updated data were included in the final analysis on survival and perioperative outcomes (Table 2). Overall, a total of 3749 patients who underwent EPP for MPM were identified from the 58 selected studies, with 2462 patients included in the final evaluation.
Verbal skills were assessed at approximately ages 2, 3, 5, and 9 years for 206 children with a clinical diagnosis of autism (n = 98), pervasive developmental disorders-not otherwise specified (PDD-NOS; n = 58), or nonspectrum developmental disabilities (n = 50). Growth curve analyses were used to analyze verbal skills trajectories over time. Nonverbal IQ and joint attention emerged as strong positive predictors of verbal outcome. The gap between the autism and other 2 groups widened with time as the latter improved at a higher rate. However, there was considerable variability within diagnostic groups. Children with autism most at risk for more serious language impairments later in life can be identified with considerable accuracy at a very young age, while improvement can range from minimal to dramatic. (PsycInfo Database Record (c) 2022 APA, all rights reserved)
Over the past 20 years a variety of treatments have been developed to remediate deficits associated with autism. Since the early 1990s, Social Stories™ have been suggested to positively affect the social development of children with autism spectrum disorder (ASD). Despite much research, there remains uncertainty regarding the effectiveness of this modality. We conducted a systematic review of the literature using pre-defined, rigorous methods. Studies were considered eligible if they were controlled trials evaluating Social Stories™ among persons with ASD. Two reviewers independently screened articles for inclusion, applied eligibility criteria, extracted data, and assessed methodological quality. A qualitative analysis was conducted on six eligible controlled trials. Five of the six trials showed statistically significant benefits for a variety of outcomes related to social interaction. This review underscores the need for further rigorous research and highlights some outstanding questions regarding maintenance and generalization of the benefits of Social Stories™.
The genetic locus encoding KIBRA, a member of the WWC family of proteins, has recently been shown to be associated with human memory performance through genome-wide single nucleotide polymorphism screening. Gene expression analysis and a variety of functional studies have further indicated that such a role is biologically plausible for KIBRA. Here, we review the existing literature, illustrate connections between the different lines of evidence, and derive models based on KIBRA’s function(s) in the brain that can be further tested experimentally.
Introduction
To fully understand how the acquisition, maintenance and recall of memories is achieved is one of the holy grails of neuroscience. Since the definition of the g-factor for “general cognitive ability” over a century ago (Spearman, 1904) and the characterization of the landmark patient H.M. in 1957 (Scoville and Milner, 1957), the field of cognitive neuropsychology has been attempting to elucidate the brain structures and molecular players involved in these processes. However, most of this progress has been reached stepwise, working forward from brain structure to structure and molecule to molecule in a hypothesis-driven fashion (Lee and Silva, 2009). Recently, the advent of high-throughput genomic polymorphism scanning technologies has revolutionized our ability to search for the common genetic drivers of memory and other cognitive domains in a hypothesis-free manner (Goldberg and Weinberger, 2004; Fisher, 2006; Payton, 2006, 2009; Potkin et al., 2009). In the case of KIBRA (also known as WWC1 for WW and C2 domain containing 1), the topic of this review, evidence for a link to cognition occurred independently both from genetic association studies (Papassotiropoulos et al., 2006) and from biochemical work (Büther et al., 2004).
Beyond understanding the way memory works, one practical goal of studying genes with relation to cognitive performance is to identify new treatment strategies that cope with the various cognitive disabilities for which treatment options are urgently needed. The population aging in industrialized countries is expected to result in a strong increase in cognitive health problems in the next decades, both as a result of aging-associated cognitive decline, as well as increased prevalence of neuropsychiatric diseases such as Alzheimer’s disease. Here, novel discoveries in cognition are sought to broaden the spectrum of pharmacologically addressable targets and hypotheses. The KIBRA-pathway appears to present a novel promising inroad for this, since KIBRA has been both linked to normal cognitive performance and to Alzheimer’s disease.
Here, we highlight the most important functional aspects of KIBRA, and connect genetic and biochemical data to aid in the formation of hypotheses related to KIBRA’s cellular and molecular function.
The KIBRA Gene and Protein
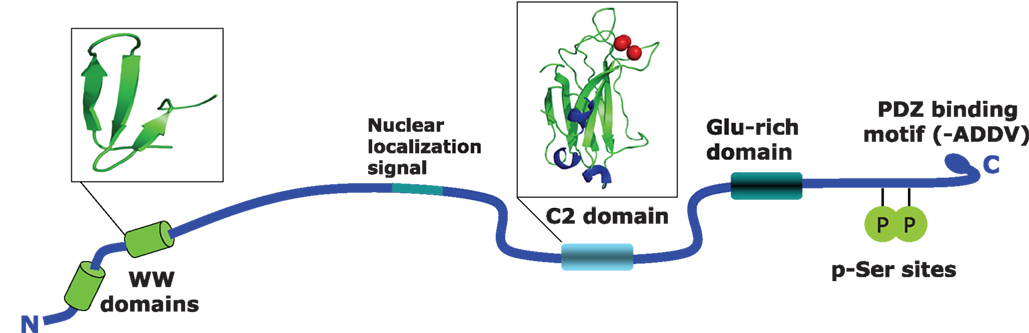
KIBRA was first cloned and initially characterized by Kremerskothen et al. (2003) as a molecule that interacts with the postsynaptic protein dendrin. Human KIBRA, encoded by the WWC1 gene, is located on chromosome 5q35.1, and contains 1113 amino acids and has a predicted size or 125.3 kDa. From N- to C-terminus a number of known protein domains have been identified which are depicted in Figure 1. The WW-domains (aa 6–86), which cover a stretch of 35–40 amino acids contain two conserved tryptophan residues. These domains are responsible for the interaction with various proteins containing proline-rich sequences (PPxY). A putative nuclear localization signal was identified between amino acids 361–376 (Rayala et al., 2006). A C2 domain composed of two four-stranded β-sheets is located between amino acid 655 and 783. The 130 residues of the C2 domain are involved in binding phospholipids in a calcium-dependent manner. C2 domains are found in proteins with functions ranging from signal transduction to vesicular trafficking (Rizo and Sudhof, 1998). Calcium binding induces a change in the electrostatic potential, which enhances phospholipid binding. A glutamic acid-rich region is located between amino acids 845 and 873 (Kremerskothen et al., 2003; Rayala et al., 2006). Lastly, a putative class III PDZ-binding sequence has been identified between amino acids 1110 and 1113 (Duning et al., 2008).

[View Larger Version of this Image]
Figure 1. Structural features of the human KIBRA protein. Shown are the identified domains in the KIBRA protein. WW domains are located between position 6 and 86, and cover about 40 amino acids containing two conserved tryptophan residues. WW domains are generally thought to be responsible for the interaction with various proteins that contain proline-rich sequences such as PPxY. The C2 domain is located between amino acids 655 and 783. This is a conserved membrane targeting motif composed of β-sheets. The 130 residues of C2 are involved in binding phospholipids in a calcium-dependent manner. The PKCζ binding region is located at amino acids 953–996 and contains two serine residues that can be phosphorylated by the kinase. The last four amino acids contain a PDZ binding motif.
Human Genetic Evidence for KIBRA’s Role in Cognition
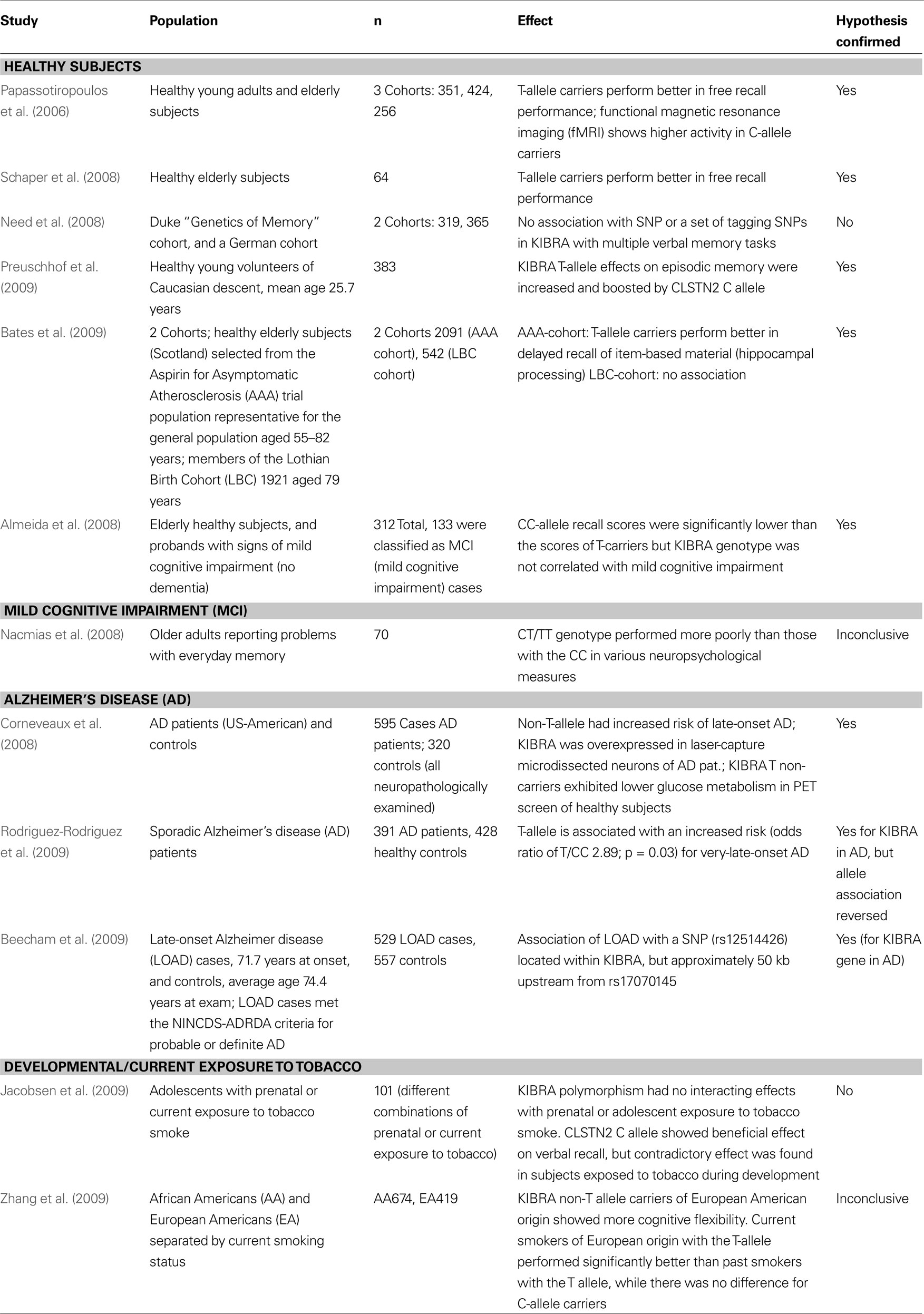
KIBRA has come into the focus of the neurogenetics field following the publication of human evidence pointing to an involvement of the gene in memory performance and cognition (Papassotiropoulos et al., 2006). In that publication, the authors report that carriers of the KIBRA/ WWC1 (rs17070145) T allele or, to a lesser extent, the calsyntenin 2 (CLSTN2) rs6439886 T allele performed significantly better on multiple episodic memory tasks than those homozygous for the C allele at either polymorphism. Furthermore, using functional magnetic resonance imaging (fMRI), they observed that brain activation (measured as oxygen extraction from blood) in key areas associated with memory retrieval was significantly greater in a selection of 15 WWC1 (rs17070145) T-allele-noncarriers than in 15 T-allele-carriers during an episodic memory task. After this initial finding, a considerable number of studies examined the WWC1 polymorphism in different contexts of cognition and in different populations (summarized in Table 1 and reviewed below).

[View Larger Version of this Table]
Table 1. Summary of the genetic association data available on the KIBRA polymorphism and memory and Alzheimer’s disease. Twelve studies that examine the KIBRA polymorphism in different populations are listed that were published before December 2009. Summarized are the citation and publication year, the population(s) examined, the study size, the effect(s) observed, and fi nally whether the study confi rmed the association of the rs17070145 SNP with memory performance.
A number of studies have examined the role of the rs17070145 polymorphism on the cognitive performance of non-demented individuals. The original finding was first confirmed in a small German cohort (n = 64) of healthy elderly individuals with a mean age of approximately 67 (Schaper et al., 2008). A second study also confirmed the influence of the T-allele on memory performance in a cohort of 312 elderly individuals between the ages of 50–89 (Almeida et al., 2008). Of note, 136 individuals in their study met diagnostic criteria for mild cognitive impairment (MCI) and they were able to identify an excess risk for MCI among non-T-allele-carriers, although it failed to reach statistical significance. A study on individuals with subjective memory complaints found that family history of dementia was significantly elevated in KIBRA non-T-allele-carriers while these same individuals exhibited better performance on verbal episodic memory tests (Nacmias et al., 2008).
In mid-2008 a report of non-replication of the influence of rs17070145 on episodic memory was published (Need et al., 2008). In this study the authors utilized two cohorts of European genetic origin. The first cohort (n = 300) was examined using a verbal recall memory test and the second cohort (n = 365) was phenotyped with the exact same auditory verbal learning test (AVLT) that was used in the original publication (Papassotiropoulos et al., 2006). The authors failed to see any association of rs17070145 with memory performance in either cohort although they reported to be well powered to do so. Additionally, some tagging SNPs within the KIBRA genetic locus were tested and no significant association signals could be identified following multiple testing corrections. In the concluding paragraph of this work the authors indicate that their non-replication findings are likely a good indicator of the challenges facing the study of the genetics of human memory and specifically point to the need for consistency in phenotyping across any replication efforts.
Finally, the largest replication effort was published in 2009 (Bates et al., 2009). In two cohorts numbering over 2,500 individuals combined, association between rs17070145 and memory performance was confirmed. Using the AVLT and the Wechsler Logical Memory Test, it was possible to demonstrate an association between rs17070145 T-allele-carrier status and item-distinctiveness storage and/or recollection. The authors suggest that KIBRA action is involved primarily in the conscious recall of item-based information and underscore the importance of attention to phenotyping when attempting to replicate a genetic association. The most recent study in healthy individuals replicated the positive effects of the KIBRA T-allele on episodic memory in a cohort of 383 individuals and showed that the effect size of the KIBRA polymorphism was modulated by the associative components of the task (Preuschhof et al., 2009). Additionally, this work suggests an interaction between KIBRA and CLSTN2 in episodic memory as the effect of carrying the rs17070145 T-allele was enhanced in carriers of the CLSTN2 rs6439886 C-allele.
Recently, KIBRA was also examined in the context of developmental or current exposure to nicotine (Jacobsen et al., 2009; Zhang et al., 2009). One study assessed association of SNP rs17070145 with verbal and visuospatial memory and fMRI changes in adolescents and could not find any significant influence of the polymorphisms either alone or in interaction with smoking habits or smoking exposure (Jacobsen et al., 2009). However, this study only examined 101 subjects with various combinations of prenatal and current nicotine exposure. The most recent study analyzed cognitive flexibility measures with the Wisconsin Card Sorting Test, ethnicity, and smoking habits in relation to the rs17070145 polymorphism (Zhang et al., 2009). In European Americans homozygous for the C-allele, an association with better cognitive flexibility was found. Curiously, current smokers of European origin with the T-allele performed significantly better than past smokers with the T-allele while there was no difference for C-allele carriers. There was no difference in performance in subjects of African American ethnicity. Association of the C instead of the T allele with better cognitive performance in different studies may be caused by differential effects of the polymorphism itself or yet unknown functional polymorphisms in the KIBRA gene in linkage disequilibrium with SNP rs17070145 on different domains of cognitive performance (e.g., more frontally located functions versus hippocampal functions).
KIBRA and Alzheimer’s Disease: Clues from the Genome and Transcriptome
Based on the associations with memory performance in healthy subjects, the link between KIBRA and Alzheimer’s disease was also examined. One study demonstrated that the rs17070145 C-allele was significantly associated with increased risk for developing late-onset AD (Corneveaux et al., 2008). However, another study showed that the T-allele was associated with an increased risk for very-late-onset AD (Rodriguez-Rodriguez et al., 2009). These differential findings may be reflective of differences in the genetic background of the studied samples, in the case/control selection criteria, or simply in the statistical power differences between the studies. A study in early 2009 also noted an association between KIBRA and late-onset AD risk (Beecham et al., 2009). Utilizing imputation to compare their data set generated on the Illumina HumanHap beadchip with data generated on a similar density Affymetrix array in a large AD study of 1,411 cases and controls (Reiman et al., 2007), the authors reported on a SNP located within KIBRA (rs12514426) that was associated [joint analysis p-value (uncorrected) = 0.000928] with late-onset AD. This SNP is in very low hypothetical linkage disequilibrium with rs17070145, but nonetheless represents a third independently reported genetic association between KIBRA and late-onset AD. Of note, the same analysis also yielded several hits in the SORL1 locus, a gene with strong biological and genetic links to Alzheimer’s disease (Andersen et al., 2005; Rogaeva et al., 2007; Dodson et al., 2008; Mayeux and Hyslop, 2008; Ma et al., 2009).
Two additional pieces of experimental evidence link KIBRA to AD (Corneveaux et al., 2008). First, the expression of KIBRA itself and the genes encoding several of its direct binding partners (DYNLL1, PRKCZ, SNX4, and KIAA0513) are significantly altered in non-tangle bearing neurons in key brain regions in AD patients versus matched controls. In neurons laser-dissected from the hippocampus, middle temporal gyrus, and posterior cingulate, KIBRA mRNA was found to be significantly increased between 2.4- and 3.0-fold, while in those same brain regions PKCζ mRNA was decreased by 2.3- to 3.3-fold. No significant changes were observed in the primary visual cortex, a region that is largely spared by AD pathology. Interestingly, only significant changes in PKCζ mRNA were detected in the hippocampus and middle temporal gyrus from patients with MCI (Huentelman lab communication, unpublished results), suggesting that perhaps the transcriptional alteration of these molecular species may follow a temporal pattern that could be related to dementia progression. Secondly, a link between rs17070145 T-allele carrier status and brain hypometabolism of fluorodeoxyglucose (FDG) as measured using positron emission tomography was described. It was shown that significant decreases in FDG uptake were noted in the precuneus and posterior/mid cingulate regions of neurologically normal individuals who were at elevated risk for developing AD. Some of these regions overlapped with previously identified hypometabolic changes in individuals stratified by APOE E4 carrier status (Reiman et al., 2004; Buckner et al., 2005; Reiman et al., 2005).
In conclusion, several pieces of evidence point to a link between KIBRA and Alzheimer’s disease. At present, the genetic link appears not as clear-cut as with memory performance in healthy individuals and therefore requires further replication. An interesting question is whether this link represents an extension or consequence of the involvement of KIBRA in episodic memory performance or possibly that it represents an independent function of KIBRA in the pathophysiology of Alzheimer’s disease.
Expression Patterns of KIBRA mRNA and Protein
KIBRA is predominately expressed in the kidney and brain in the adult organism. In the rodent and human brain, KIBRA is expressed in memory-related structures (including the hippocampus and cortex), the cerebellum, and in the hypothalamus (Johannsen et al., 2008). During brain development, expression has been shown to decrease from juvenile postnatal stages to the adult animal (Johannsen et al., 2008). On a subcellular level within neurons, KIBRA shows a somatodendritic staining pattern with enrichment in the perinuclear region and in postsynaptic structures. In subcellular fractionation experiments, KIBRA was heavily enriched in the postsynaptic density (PSD) fraction (Johannsen et al., 2008). However, nuclear localization has also been noted (Rayala et al., 2006). So far, it is unclear if different localizations are due to the type of cells investigated, experimental conditions, or possible modified variants of KIBRA.
Interacting Partners of KIBRA
KIBRA has been shown to participate in a number of cellular functions such as cell polarity and migration, vesicle transport, transcriptional regulation and synaptogenesis. Most hints to its function have come from the study of interaction partners identified in yeast two hybrid screens. To date, 10 direct interaction partners have been described (Figure 2). The two WW-domains, which cover a stretch of 35–40 amino acids containing two conserved tryptophan residues, appear responsible for many of the interactions with the identified proteins. The first interaction partner of KIBRA, identified by a yeast two hybrid screen, was the postsynaptic protein dendrin (KIAA0749) (Kremerskothen et al., 2003). Dendrin was first characterized as a dendritic protein whose expression is altered by sleep deprivation in the forebrain of rats (Neuner-Jehle et al., 1996). Dendrin is translated from a dendritically localized mRNA and interacts with alpha-actinin and the synaptic scaffolding molecule S-SCAM (Kremerskothen et al., 2006).
Individuals with autism spectrum disorders experience pervasive social and communication skill deficits. One of the most popular treatment recommendations to address these deficits is the use of social stories. Although social stories are beneficial at reducing many disruptive behaviors, empirical evidence for their use to increase social and communication behaviors is limited. The present study, conducted in a natural setting, investigated the use of social stories to increase the prosocial behaviors of verbal initiations and contingent responses in students with either autism or Asperger\'s disorder. Results suggested that the efficaciousness of this widely accepted intervention is questionable and in need of further research.
Subcortical brain structures are integral to motion, consciousness, emotions and learning. We identified common genetic variation related to the volumes of the nucleus accumbens, amygdala, brainstem, caudate nucleus, globus pallidus, putamen and thalamus, using genome-wide association analyses in almost 40,000 individuals from CHARGE, ENIGMA and UK Biobank. We show that variability in subcortical volumes is heritable, and identify 48 significantly associated loci (40 novel at the time of analysis). Annotation of these loci by utilizing gene expression, methylation and neuropathological data identified 199 genes putatively implicated in neurodevelopment, synaptic signaling, axonal transport, apoptosis, inflammation/infection and susceptibility to neurological disorders. This set of genes is significantly enriched for Drosophila orthologs associated with neurodevelopmental phenotypes, suggesting evolutionarily conserved mechanisms. Our findings uncover novel biology and potential drug targets underlying brain development and disease.
Subcortical brain structures are essential for the control of autonomic and sensorimotor functions1,2, the modulation of processes involved in learning, memory and decision-making3,4, and in emotional reactivity5,6 and consciousness7. They often act through networks influencing input to and output from the cerebral cortex8,9. The pathology of many cognitive, psychiatric and movement disorders is restricted to, begins in or predominantly involves subcortical brain structures and related circuitries10. For instance, tau pathology has shown to manifest itself early in the brainstem of individuals with Alzheimer’s disease before spreading to cortical areas through efferent networks11. Similarly, the formation of Lewy bodies and Lewy neurites in Parkinson’s disease appears early in the lower brainstem (and olfactory structures) before affecting the substantia nigra12.
Recent investigations have identified genetic loci influencing the volumes of the putamen, caudate and pallidum, which pointed to genes controlling neurodevelopment and learning, apoptosis and the transport of metals13,14. However, a larger study combining these samples and including individuals of a broad age range across diverse studies would enable increased power to identify additional novel genetic variants contributing to variability in subcortical structures, and further improve our understanding of brain development and disease.
We sought to identify novel genetic variants influencing the volumes of seven subcortical structures (the nucleus accumbens, amygdala, caudate nucleus, putamen, globus pallidus, thalamus and brainstem (including the mesencephalon, pons and medulla oblongata)), through genome-wide association (GWA) analyses in almost 40,000 individuals from 53 study samples (Supplementary Tables 1-3) from the Cohorts of Heart and Aging Research in Genomic Epidemiology (CHARGE) consortium, the Enhancing Neuro Imaging Genetics through Meta-Analysis (ENIGMA) consortium and UK Biobank.
Go to:
Results
Heritability.
To examine the extent to which genetic variation accounts for variation in subcortical brain volumes, we estimated their heritability in two family-based cohorts: the Framingham Heart Study (FHS) and the Austrian Stroke Prevention Study Family Study (ASPS-Fam). Our analyses were in line with previous studies conducted in twins15, suggesting that variability in subcortical volumes is moderately to highly heritable. The structures with the highest heritability in the FHS and ASPS-Fam were the brainstem (ranging from 79–86%), caudate nucleus (71–85%), putamen (71–79%) and nucleus accumbens (66%), followed by the globus pallidus (55–60%), thalamus (47–54%) and amygdala (34–59%) (Fig. 1 and Supplementary Table 4). We additionally estimated single-nucleotide polymorphism (SNP)-based heritability (h2g) using genome-wide complex trait analysis (GCTA) in the Rotterdam Study, and linkage disequilibrium score regression (LDSC) in the full European sample. As expected, SNP-based heritability estimates were somewhat lower, ranging from 17% for the amygdala to 47% for the thalamus using GCTA, and ranging from 9% for the amygdala to 33% for the brainstem using LDSC. These values are consistent with heritability estimates reported by UK Biobank14.
Besides the direct economic and social burden of myopia, associated ocular complications may lead to substantial visual loss. In several population and clinic-based cohorts, case–control and cross-sectional studies, higher risks of posterior subcapsular cataract, cortical and nuclear cataract in myopic patients were reported. Patients with high myopia (spherical equivalent at least –6.0 D) are more susceptible to ocular abnormalities. The prevalent risks of glaucoma were higher in myopic adults, and risks of chorioretinal abnormalities such as retinal detachment, chorioretinal atrophy and lacquer cracks increased with severity of myopia and greater axial length. Myopic adults were more likely to have tilted, rotated, and larger discs as well as other optic disc abnormalities. Often, these studies support possible associations between myopia and specific ocular complications, but we cannot infer causality because of limitations in study methodology. The detection and treatment of possible pathological ocular complications is essential in the management of high myopia. The ocular risks associated with myopia should not be underestimated and there is a public health need to prevent the onset or progression of myopia.
Myopia has reached epidemic proportions and is already a large public health problem in certain parts of the world, including East Asia (Grosvenor, 2003). The rates of high myopia, and possibly pathological myopia, appear to be rising in Asia and other parts of the world. Cross-sectional prevalence surveys across the United States (National Health and Nutrition Examination Survey, Baltimore Eye Survey, Beaver Dam Eye Study, and the Framingham Offspring Eye Study) suggest that there are higher myopia prevalence rates with more recent birth cohorts (Sperduto et al., 1983; Wang et al., 1994; The Framingham Offspring Eye Study Group, 1996; Katz et al., 1997). The evidence for this phenomenon from repeated prevalence surveys, however, needs to be further substantiated because of differences in methodology (Lin et al., 1988,1999). The apparent worldwide rise in the prevalence of myopia has a large public health impact because of the associated concomitant increase in potentially blinding ocular conditions.
Common definitions of high myopia or myopia with increased risks of ocular morbidity, include spherical equivalent (SphE) of at least –6.0 D, SphE at least –8.0 D, or SphE at least –10.0 D. Ocular pathology is usually due to excessive elongation of the eyeball and associated with pathological changes in the fundus (Goldschmidt, 1988; Tokoro, 1988). The terms ‘malignant myopia’, ‘degenerative myopia’ and ‘pathological myopia’ have also variously been used to describe myopia accompanied by degenerative changes in the sclera, choroid, retinal pigment epithelium and associated compromises in visual function (Duke-Elder, 1970; Daubs, 1982). Tokoro defined pathological myopia as myopia caused by pathological axial elongation and showed that the prevalence increased from 0.5% in junior high school Japanese students to a peak of approximately 3% in 29-year-old adults (Tokoro, 1988).
We aim in this review to summarize the best evidence of myopia-associated ocular pathologies, including cataract, glaucoma, chorioretinal abnormalities, optic disc abnormalities and age-related macular degeneration. Acronyms are summarized
The “old school” and the “new school” of narratology seem to share a referential
bias in their view of fictional minds. (1) Classical studies assume that via a metic-
ulous differentiation of voices and viewpoints, we are able to extract a “reliable”
representation of the fictional world. (2) Recent cognitive approaches tend to
regard fictional and actual minds as being based on precisely the same cognitive
schemata; we should ask ourselves, however, whether the natural parameters of
thought and those of fiction actually converge. In this paper, I wish to demon-
strate the challenge that fictional characters representing each other’s perceptions,
thoughts and feelings issue to both classical and cognitive approaches to fiction-
al minds. I suggest that fictional agents – not only narrators but focalizers as
well – may take advantage of precisely the same techniques of constructing the
minds of others as are used in omniscient narration. Furthermore, I try to dem-
onstrate how this feature of literary representation inherently problematizes the
“naturalizing” of (fictional) mental functioning. I will start with examples from
homodiegetic narration (Ford Madox Ford, Richard Ford) and then widen the
scope to more problematic instances in the heterodiegetic mode (Emmanuèle
Bernheim, Fay Weldon). I argue that the “narrative tendency” of a character con-
structing other characters’ minds potentially mitigates the binarity of homodi-
egetic and heterodiegetic narration. Finally, such speculating and narrativizing
– and perhaps fantasizing, hallucinating – fictional minds provide a parallel, not
In this paper, I wish to avoid such referential biases and concentrate on the
peculiarly novelistic ways of thematizing human consciousness. Methodologi-
cally, however, I try to draw on both the classical formalist approaches and the
cognitive-based innovations whenever they seem to be sensitive enough to the
literary phenomena in question. My particular aim is to highlight the functions
of free indirect discourse (and related techniques of rendering fictional mental
activity) in embedded or recursive representation of consciousness. The problem
will be illustrated with four exemplary texts: novels by Ford Madox Ford (The
Good Soldier, 1915) and Emmanuèle Bernheim (Sa femme/Sa Femme, Or the
Other Woman, 1993/1994) and short stories by Richard Ford (‘Privacy,’ 2001)
and Fay Weldon (‘Weekend,’ 1978).
In the field of cognitive science, it was Daniel Dennett who first, in 1983, dis-
cussed the levels of intentionality as a cognitive model: mental “worlds” can be
represented as a recursive system that is potentially infinite as in statements like
“X thinks that Y believes A to be mad at B” and so on (see, eg. Zunshine 2003:
278). Such embedded states of mind occur frequently in novels concentrating on
the psychological processes of the characters – however, it is the peculiar nature
234Maria Mäkelä
of novelistic presentation that it often avoids such overt marks of intentional-
ity as explicit mentions of who is behind each represented thought, opinion, or
discourse. This ambiguity of voices/subjectivities has been a perennial topic in
classical narratology, but, as indicated above, the main cause for the concern has
been the ambiguity between objective narratorial report and the subjective ele-
ments attributable to a fictional character (e.g. Pascal 1977). A far less scoured
area of novelistic consciousness representation has to do with characters repre-
senting each other’s perceptions, thoughts and feelings; a glance at the brand new
Routledge Encyclopedia of narrative theory attests to the absence of research on
this particular area (see, eg. the entry on THOUGHT AND CONSCIOUSNESS
REPRESENTATION by Palmer in Herman & Jahn & Ryan (ed.) 2005: 602–
607). What I would like to suggest in this paper is that fictional agents – not only
narrators but focalizers as well – may take advantage of precisely the same tech-
niques of constructing the minds of others as are used in omniscient narration.
Furthermore, I try to demonstrate how this feature of literary representation
inherently problematizes the “naturalizing” of (fictional) mental functioning.
Obviously, the potential of embedded consciousness representation has not
passed without any notice among narratologists. Moshe Ron provides examples
from The Good Soldier (Ron 1981: 23–24) as well as from Henry Miller’s Sexus
(ibid.: 31), pointing out the narrative possibilities of “focalizers focalizing each
other” and the challenge this kind of representation issues to the “Mimetic Lan-
guage Game.” Brian McHale (1983: 33–34) also singles out this kind of represen-
tation in Ulysses (is Bloom constructing Gerty MacDowell’s point of view within
his own or does the narration offer a “real” shift to Gerty’s mind?) to prove that,
unlike Banfield argues, we definitely need contextual cues – in addition to lin-
guistic ones – in order to attribute represented thought segments to individual
characters. Yet, even the cognitively inclined theorists who (briefly) touch upon
cases of embedded consciousness representation (Jahn 1992: 357–358; Zunshine
2003: 278–281) consider its thematic implications only in relation to the assumed
“what is really going on in this passage” instead of paying attention to the chal-
lenge such representation issues to referentiality, experientiality (cf. Fludernik
1996) – and to cognitive narratology, for that matter.
In the following, I try to demonstrate how fictional mental worlds embed-
ded within each other challenge both classical and cognitive approaches to
fictional minds. I will start with examples from homodiegetic narration (Ford
235Possible minds
Madox Ford and Richard Ford) and then widen the scope to more problematic
instances in the heterodiegetic mode (Bernheim, Weldon); finally, I argue that
embedded consciousness representation potentially mitigates the binarity of
first-person (homodiegetic) and third-person (heterodiegetic) narration (a nar-
ratological idée reçue). The “narrative tendency” of a character constructing the
other characters’ minds conflicts both 1) with the (natural narratology) logic
that epistemic restrictions result in narrative restrictions as well; and 2) with the
idea that in a heterodiegetic narrative situation, all diegetic material should be
attributed to the narrator while the role of the character is restricted to “reveal-
ing” him/herself through the mimetic glimpses the narration offers to the read-
er. Furthermore, novels with such speculating and narrativizing – and perhaps
fantasizing, hallucinating – characters are also self-reflective in that they reflect
both the novelist’s and the reader’s process of interpretation and construction of
fictional worlds and consciousnesses. The analogy between embedded mental
spaces and fictional world construction will also lead us – at least for a while
– to the cognitive linguistic realms of Possible Worlds Theory and Text World
Theory (cf. Ryan 1991; Doležel 1998; Gavins 2003).
Instead of pledging allegiance to the linguistic or otherwise strict definitions
of free indirect discourse, I attempt to use this classical concept as representing
the narrative “border phenomena,” not just between indirect and direct repre-
sentation, but between the mental spaces (possible worlds and possible minds)
evoked in narration. Consequently, the “freedom” of these narrative situations
refers to the implicitness of the levels of intentionality and the “indirectness”
to all the mediating agents that separate the reader from the referred subject of
representation – the last mental domain at the end of the narrative food chain.
The oxytocin receptor gene (OXTR) has been studied as a risk factor for autism spectrum disorder (ASD) owing to converging evidence from multiple levels of analysis that oxytocin (OXT) has an important role in the regulation of affiliative behavior and social bonding in both nonhuman mammals and humans. Inconsistency in the effect sizes of the OXTR variants included in association studies render it unclear whether OXTR is truly associated with ASD, and, if so, which OXTR single-nucleotide polymorphisms (SNPs) are associated. Thus, a meta-analytic review of extant studies is needed to determine whether OXTR shows association with ASD, and to elucidate which specific SNPs have a significant effect on ASD. The current meta-analysis of 16 OXTR SNPs included 3941 individuals with ASD from 11 independent samples, although analyses of each individual SNP included a subset of this total. We found significant associations between ASD and the SNPs rs7632287, rs237887, rs2268491 and rs2254298. OXTR was also significantly associated with ASD in a gene-based test. The current meta-analysis is the largest and most comprehensive investigation of the association of OXTR with ASD and the findings suggest directions for future studies of the etiology of ASD.

 www.nature.com
www.nature.com
There is emerging evidence implicating oxytocin receptor gene (OXTR) DNA methylation (DNAm) in social behaviour. This review investigated its association with autism spectrum disorder (ASD) characteristics and related social dimensions, both in individuals with and without ASD. Twelve articles investigating OXTR DNAm in relation to ASD, social perception/cognition and social anxiety were included. We found that hypermethylation is associated with (i) higher quantitative autism traits in adults, reflecting a higher incidence of autism characteristics, (ii) increased brain activity while performing social tasks (indicating a higher need for resources) and (iii) decreased functional connectivity. (iv) Contradictory, hypomethylation was found to be present in children (especially boys) with ASD and was also associated with more social anxiety. While the included studies displayed a large variability, for example in terms of population characteristics, analysed OXTR DNAm regions, and adopted scales/questionnaires, an initial developmental pattern of results emerged, suggesting an association between hypermethylation of OXTR and autism traits in adults. Nonetheless, future studies are warranted to corroborate these initial conclusions.
Graphical abstract
OXTR is a single copy, located on chromosome 3 (hg38, 3:8750408-8769628). The gene contains 3 introns and 4 exons. This figure shows the structure of OXTR with the approximate location of the CpG sites -860, -901, -924, -934, -959, -982, -989 (upstream from the translation start site) and the rs53576 SNP. The red lines indicate the investigated CpG sites/regions and the green lines show studies investigating the SNP rs53576 as well, for the included studies in this review: 1. (Gregory et al., 2009), 1A: exons 1-3, introns 1 (sites -860; -901;-924; -934; -959) and 2, and 1B. intron 3; 2. (Elagoz Yuksel et al., 2016), exon 1-3 and intron 1 and 2; 3. (Andari et al., 2020), intron 1 (site -989); 4. (Siu et al., 2021), intron 1 (site -982); 5. (Jack et al., 2012), intron 1 (site -934); 6. (Puglia et al., 2015), intron 1 (site -934); 7. (Rijlaarsdam et al., 2017), exon 2 (average of 3 sites); 8. (Puglia et al., 2018), intron 1 (site -934); 9. (Chen et al., 2019), intron 1 (sites -934, -924 and -901); 10. (Krol et al., 2019), intron 1 (site -924); 11. (Chagnon et al., 2015), exon 3; 12. (Ziegler et al., 2015), exon 3 (average over 12 sites). * The arrows schematically show OXTR hyper- (↑) or hypomethylation (↓) in ASD or in association with autism traits, social perception/cognition or social anxiety per study.
The diagnostic histories of autism, dyslexia,
and hyperlexia are complex. Because these
conditions share both convergent and diver-
gent properties, it is important to understand
these relationships, especially in the case of
research and how we interpret bodies of work
which span decades of fluctuating criteria. It is
also important to synthesize what we already
know about the morphology of these condi-
tions and pinpoint what we still don’t. Autism
and dyslexia, for instance, share antipodal
cerebral morphologies, such as minicolumnar
density, neuropil width, cell size, corpus callo-
sal volume, gyral complexity, gyral window
size, and cerebral volume, while hyperlexia
has not been studied in this fashion, although it
shares much in common with autism. Mean-
while, the fluctuation in criteria of dyslexia

Hyperlexia is a condition rarely reported in Spanish-speaking children, characterized by the ability to recognize written words without formal training. We present two unrelated autistic children with hyperlexia from Colombia (South America) who were followed for 8 years with formal neuropsychological evaluations of language, motor skills, visual perception, attention and behavior. Both children taught themselves to read before 5 years of age but showed minimal comprehension; both displayed obsessional reading and difficulties in social skills and attention. Brain CT scans were normal. Hyperlexia has been associated with hyperactivation of the left superior temporal cortex; we conclude that the orthographic route is a probable mechanism for the development of hyperlexia.
Shovel shape of upper incisors is a common characteristic in Asian and Native American populations but is rare or absent in African and
European populations. Like other common dental traits, genetic polymorphisms involved in the tooth shoveling have not yet been
clarified. In ectodysplasin A receptor (EDAR), where dysfunctional mutations cause hypohidrotic ectodermal dysplasia, there is a nonsy-
nonymous-derived variant, 1540C (rs3827760), that has a geographic distribution similar to that of the tooth shoveling. This allele
has been recently reported to be associated with Asian-specific hair thickness. We aimed to clarify whether EDAR 1540C is also associated
with dental morphology. For this purpose, we measured crown diameters and tooth-shoveling grades and analyzed the correlations
between the dental traits and EDAR genotypes in two Japanese populations, inhabitants around Tokyo and in Sakishima Islands. The
number of EDAR 1540C alleles in an individual was strongly correlated with the tooth-shoveling grade (p ¼ 7.7 3 1010 ). The effect
of the allele was additive and explained 18.9% of the total variance in the shoveling grade, which corresponds to about one-fourth
of the heritability of the trait reported previously. For data reduction of individual-level metric data, we applied a principal-component
analysis, which yielded PC1-4, corresponding to four patterns of tooth size; this result implies that multiple factors are involved in dental
morphology. The 1540C allele also significantly affected PC1 (p ¼ 4.9 3 103
), which denotes overall tooth size, and PC2 (p ¼ 2.6 3
103
), which denotes the ratio of mesiodistal diameter to buccolingual diameter.
Teeth display variations among individuals in the size and
the shape of cusps, ridges, grooves, and roots. In addition,
certain dental characteristics are predominant in certain
populations. 1,2 In Asian and Asia-derived populations,
dental variations have often been described as ‘‘Sino-
donty’’ and ‘‘Sundadonty.’’ Sinodonty, common among
East Asian and Native American populations, is a combina-
tion of dental characteristics that relatively often include
upper first and second incisors (UI1 and UI2) that are
shovel-shaped and not aligned with the other teeth,
upper first premolars (UP1) with one root, and lower
first molars (LM1) with three roots. 3 In particular, the
prevalence of tooth shoveling shows marked regional
differences, which is frequent in Asia, with a south-to-
north increasing cline, but rare or absent in Africa and
Europe. 4,5 Tooth shoveling has thus received much
attention from anthropologists as an indicator of relation-
ships among populations. The well-known contrast
between ancient and modern Asians is that remains of
the Jomon people, who were the indigenous Japanese
12,000–2,000 years ago, have a smaller tooth size and
a lower tooth-shoveling grade than modern Japanese. 1,6–8
In addition, the continuity of shovel-shaped incisors
between Homo erectus and modern humans in East Asia
was a rationale for the multiregional evolution theory of
modern humans, 9,10 although this theory is not generally
supported at present.
Characteristics of dental morphology are thought to be
determined predominantly by genetic factors. 5 However,
genetic polymorphisms associated with common dental
traits have not yet been elucidated. For tooth shoveling,
high concordance between monozygotic twins has been
observed. 11,12 On the basis of intrafamilial correlations,
three different studies have reported that the heritability
of tooth shoveling was around 0.75 in Asian and Native
American groups. 5,11,13
The recent availability of large-scale data on human
genome diversity has enabled us to screen genes highly
differentiated between populations; these make reasonable
candidate genes for population-specific phenotypes. One
nonsynonymous-derived variant predominant in East
Asian populations but absent in populations of African
and European origins is present on rs3827760, a single-
nucleotide polymorphism (SNP) in the ectodysplasin A
receptor gene (EDAR [MIM 604095]), which is also called
EDAR T1540C according to nucleotides or V370A accord-
ing to amino acids. 14–16 In humans, dysfunctional muta-
tions in EDAR as well as the ectodysplasin A gene (EDA
[MIM 300451]) are known to be responsible for hypohi-
drotic ectodermal dysplasia [MIM 129490, 224900, and
305100], a genetic disorder causing abnormal morphogen-
esis of teeth, hair, and sweat glands.17 Mouse and fish
studies have also indicated that the Eda-Edar pathway
regulates the development of organs, especially those